

Endothelial Responses to Aneurysm Initiating Hemodynamics
Our lab has implicated high wall shear stress (WSS) and positive WSS gradient (WSSG) as hemodynamic parameters involved in the initiation of intracranial aneurysms. In our studies of surgically created bifurcation in dogs and in the basilar terminus of rabbits exposed to increased flow we have shown matrix degradation and cell loss specifically localize to regions of accelerating flow where WSS is high and WSSG is positive. However, the adjacent regions experiencing equally high WSS but decelerating flow, i.e., a negative WSSG, remain undamaged, suggesting that the endothelium responds differently to positive and negative WSSG.
To elucidate the effects of high WSS and WSSG on ECs we have designed and constructed several in vitro flow loop systems. These include: (1) parallel plate flow chambers (left image) to expose ECs to uniform values of WSS, (2) a “T-chamber” which mimics impinging flow seen at bifurcations, (3) a “tapered chamber” which produces multiple WSS values in a given experiment and (4) most recently a “constant gradient chamber” (right image) which allows for the simultaneous study of high WSS, very high WSS and positive and negative WSSG.
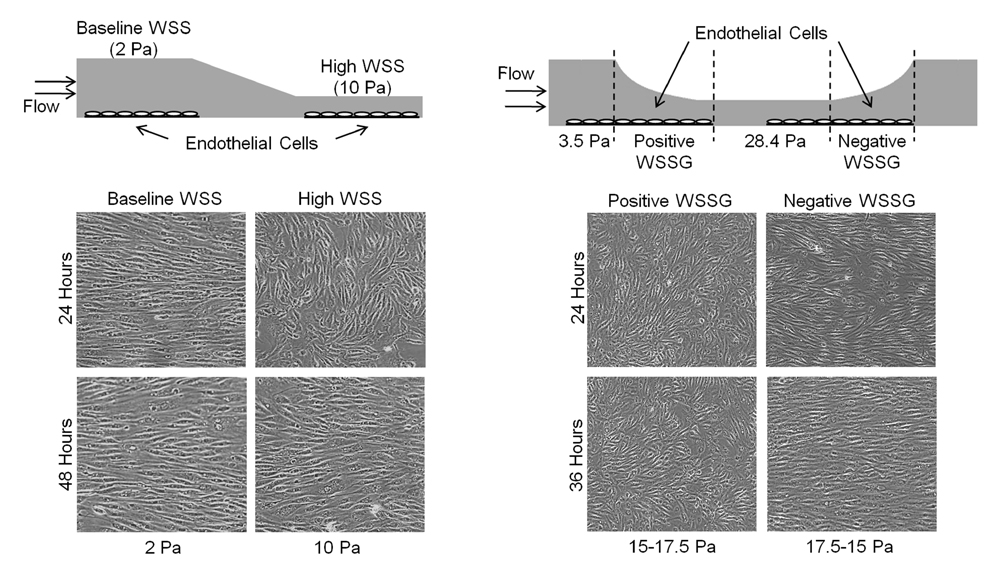
The constant gradient chamber (right image) consists of four distinct flow regions: constant-height channels to create regions of uniform WSS and converging and diverging channels to create positive and negative WSSG respectively. Using this device, we examined how fundamental EC responses - cell morphology, alignment, DNA replication, programmed cell death, and gene expression - are affected by high WSS (28.4 Pa) and positive and negative WSSG. We found that high WSS inhibited EC alignment to flow, increased EC proliferation and increased apoptosis. These responses to high WSS were either accentuated or ameliorated by WSSG: Positive WSSG (+980 Pa/m) inhibited alignment (right image) and stimulated proliferation and apoptosis, whereas negative WSSG (-1120Pa/m) promoted alignment and suppressed proliferation and apoptosis. Using the parallel chamber (left image) we found that high WSS (with no gradient) delays flow induced alignment. These results demonstrate that ECs discriminate between positive and negative WSSG under high WSS conditions.
Recognizing that EC responses to high WSS and positive WSSG may contribute to pathogenic remodeling that occurs at bifurcations preceding aneurysm formation we are also interested in identifying how ECs, under these flow conditions, alter their gene and protein expressions to signal the surrounding vessel wall to undergo maladaptive remodeling. This question is currently under investigation in the lab.
For more informtion, please contact Jennifer Dolan at jdolan(at)buffalo.edu.